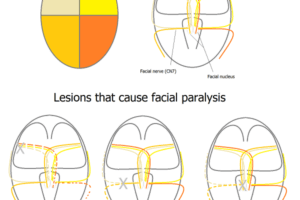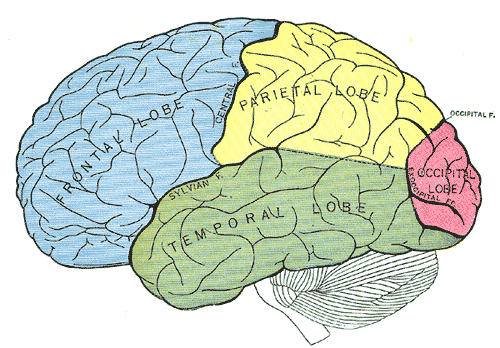
The cerebral cortex, derived from the dorsal telencephalon or pallium, consists of two hemispheres (the right and the left cerebral hemispheres), each of which is divided into five lobes.
Lobes of the Cerebral Cortex
The four lobes that are visible from a lateral view include the frontal lobe, in which “executive decisions” are made and includes motor cortical areas, the occipital lobe, devoted to vision, the parietal lobe in which several visual areas and somatosensory areas are located, and the temporal lobe, which includes areas for audition, vision, and multi-modal processing.
The insula is the fifth lobe, and is not visible from the surface of the brain. It is buried beneath the parietal and temporal lobes. The insular cortex is part of the limbic system, and is sometimes considered part of the “limbic lobe.” It’s potential roles in emotion are active areas of research.
Brodmann’s Areas
Fifty-two histologically defined regions of the cerebral cortex were identified by Korbinian Brodmann in 1909. Most of these areas have specialized functions. For example, Brodmann’s Area 17 is primary visual cortex, Area 41 is primary auditory cortex, and Area 4 is primary motor cortex. A large list of Brodmann’s Areas and a map of cortex showing their locations.
Neocortex
These external lobes are neocortex – which means “new” cortex since it is found in mammals but not in phylogentically older species. Neocortex is characterized by six layers of neurons, which when viewed in cross section appear gray, and hence is called “gray matter.” The 6 layers, or laminae, have somewhat stereotyped functions and organization. Because primary visual cortex is the most heavily studied neocortical area, the lamination of V1 is used as “typical” of sensory cortical areas.
Lamination
A. Sensory input from the thalamus enters V1 and first synapses in Layer IV (there are weaker collateral projections to Layer VI).
B. Layer IV neurons project primarily up to Layers II/III, which have both short and long range horizontal connections within cortex.
C. Layers II/III synapse onto Layer V neurons, and also onto Layer VI neurons.
D. Layers V is the main output layer of the cortex, projecting to subcortical structures like the pulvinar nucleus of the thalamus and the superior colliculus.
E. Layer VI primarily sends feedback connections to its inputs from the thalamus. It is not clear how this feedback affects the inputs to the cortex.
Lamination Differs Among Cortical Areas
While the laminar structure defined for V1 is considered “typical,” in fact lamination varies substantially across different cortical areas. For example layer IV (input layer) is tiny in motor cortex, but layer V, which sends long axonal projections into the cortico-spinal tract, is thick with large pyramidal cells.
Cell Types
Cells in the input layer (Layer IV) are primarily small, star-shaped neurons called stellate cells. Stellate cells with dendritic spines (spiny stellate cells) are glutamatergic, and therefore excitatory. Smooth stellate cells, in contrast, lack dendritic spines, use GABA as a neurotransmitter and are therefore inhibitory. Stellate cells, whether excitatory or inhibitory, are interneurons within the cerebral cortex since they project only locally.
Most of the neurons in the other layers are pyramidal cells, and are glutamatergic. Pyramidal cells project beyond the local 0.5 mm. For example, pyramidal cells in Layer II project up to 5 mm laterally within the same cortical area. Pyramidal cells in Layer V project long distances out of the cerebral cortex, either to sub-cortical structures or even into the spinal cord.
Interspersed among pyramidal cells are a variety of inhibitory interneurons, with many different morphologies.
Connections Among Areas
Cortical areas are interconnected – and these interconnections are thought to underlie the basis of cortical processing. Dogma suggests that each subsequent area processes inputs in more complex ways. There are two hypotheses of the dominant connections between areas. The classic hypothesis is that areas connect to each other through Layer II/III projection neurons whose axons enter the white matter, travel to the next area, and synapse within Layer IV. Recently this connectivity pattern has been challenged, with the alternative pattern being that areas are connected through the thalamus. For example, Layer V cells in the primary visual cortex project to the pulvinar nucleus, which in turn projects up to secondary visual area.
Cortical areas project to their matching area in the opposite hemisphere through the corpus callosum.
Archicortex and Paleocortex
More ancient cortex (allocortex) comes in the two types: paleocortex and archicortex. This type of cortex is found in mammals, but also in reptiles and avian species. Archicortex, unlike neocortex, has only three layers. The hippocampus, and associated dentate gyrus, are composed of archicortex. Paleocortex is thought to be intermediate between neocortex and archicortex. Piriform cortex (“primary olfactory cortex”) and parahippocampal gyrus are paleocortical structures.